Las proteínas de la leche se dividen en tres grupos: Las caseínas, las proteínas del lactosuero y las que forman parte de la membrana del glóbulo graso. Estas últimas representan solamente del orden del 1% del total de la proteínas de la leche
Las caseínas son las principales proteínas de la leche. Se sintetizan exclusivamente en la glándula mamaria, y en la leche se encuentran en su mayor parte formando agregados multimoleculares conocidos como “micelas de caseína”. En la leche de vaca, la caseínas representan alrededor del 80% del total de proteínas, es decir, de 25 a 28 gramos por litro de leche. En la leche humana la presencia de proteínas del lactosuero es mucho mayor, de tal forma que las caseínas son solamente del orden de la mitad de las proteínas totales, entre 5 y 8 gramos por litro.
Estructura de las moléculas de caseína
Las moléculas individuales de caseína se caracterizan en general por tener un tamaño mediano (unos 200 aminoácidos, siendo algo menor la caseína κ) contar con pocos tramos con estructura secundaria organizada, debido a la presencia de abundantes restos de prolina, y tener unidos covalentemente grupos fosfato a algunos de los restos de serina, y muy ocasionalmente a restos de treonina.
La falta de organización de las moléculas de caseína ha hecho que hasta el momento ninguna haya podido cristalizarse para llevar a cabo estudios detallados de su estructura secundaria y terciaria.
Una propiedad clásica, que ha servido durante un siglo para su definición operacional, es que las caseínas precipitan a pH 4,6, que es su punto isoeléctrico (a temperatura ambiente).
Desde el punto de vista de la estructura, en la leche bovina (y en la mayoría de las leches de otras especies) existen cuatro caseínas, conocidas como αs1, αs2, (la s del subfijo indica que son “sensibles” al calcio, es decir, que pueden precipitar al asociarse con él) β y κ. Las llamadas “caseínas γ” son simplemente fragmentos de la caseína β producidos por proteolisis por la plasmina. Todas las caseínas tienen variantes genéticas, producidas por sustitución de aminoácidos y en algunos casos por delección.
Existen diferencias en la proporción que representa cada tipo en el total de las caseínas. De entre las especies más comunes, las mayores diferencias se encuentran en el cotenido de caseína κ que representa el 3% de las caseínas de leche de búfala, el 13% de las de la leche de vaca y el 26% de las de la leche humana.
Caseína αs1
La caseína αs1 es la mayoritaria en la leche de vaca. La variante más común tiene 199 aminoácidos en su secuencia, con 8 ó 9 grupos fosfato. Desde el punto de vista estructural, está formada por tres regiones hidrofóbicas, con dos de ellas situadas en los extremos (aminoácidos 1-41, 90-113 y 132-199), y una zona muy polar (entre los aminoácidos 42 y 80), en la que se encuentran todos los grupos fosfato menos uno, lo que le da una carga neta negativa muy importante al pH de la leche (alrededor de 6,6).
La caseína αs1 de vaca contiene 17 restos de prolina, distribuidos a lo largo de toda la cadena, lo que hace que tenga muy pocas zonas con estructura secundaria organizada. La asociación con otras moléculas de caseína se produce a través de interacciones hidrofóbicas en las que está implicada fundamentalmente la zona situada entre los aminoácidos 136 y 196.
Caseína αs2
Esta caseína está formada, en la vaca, por 207 aminoácidos Se conocen varias variantes genéticas, y también varias variantes en el grado de fosforilación. La máxima fosforilación afecta a 12 serinas y una treonina. Esta caseína tiene un puente disulfuro entre las cisteínas que ocupan las posiciones 36 y 40 de la secuencia, y es más hidrofílica que la caseína αs1, con tres regiones de carga neta negativa, una de ellas en el extremo N-terminal. En la zona del extremo C-terminal se sitúan aminoácidos hidrofóbicos y con carga neta positiva.
Caseína β
La caseína β es la caseína más hidrofóbica, y presenta además estructura particular, con una clara división en dos zonas. La que corresponde al extremo C-terminal es particularmente hidrofóbica, mientras que los aminoácidos más hidrofílicos, y todos los grupos fosfato unidos a serinas, se concentran en el extremo N-terminal. La variante genética más común en la vaca está formada por 209 aminoácidos, con cinco grupos fosfato.
Estructura de la caseína β
Caseína κ
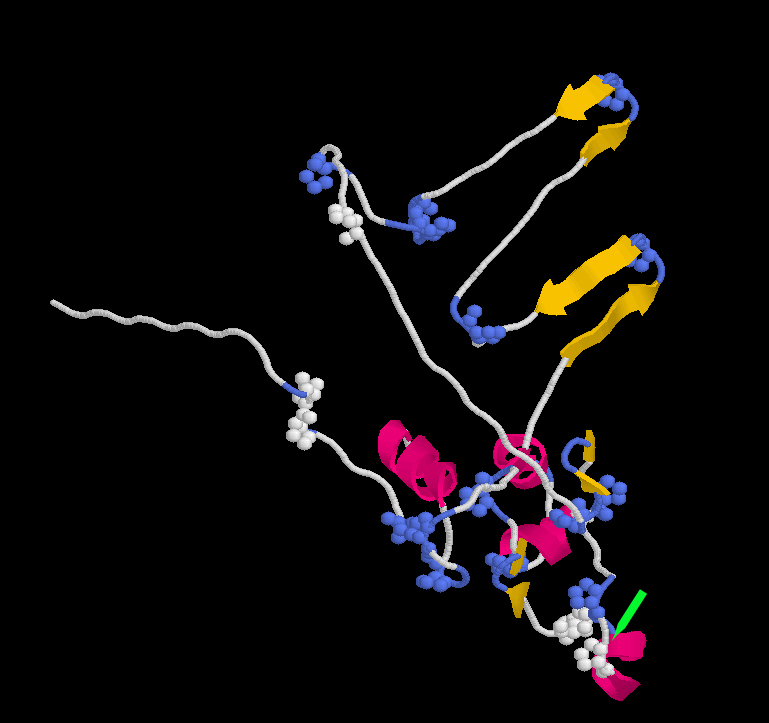
La caseína κ tiene una estructura claramente distinta de la de las otras caseínas. En primer lugar, es algo más pequeña, estando formada, en la vaca, por 169 aminoácidos. Además está muy poco fosforilada, teniendo solamente un grupo de fosfato. Esto hace que interaccione con el ión calcio mucho menos que las otras caseínas. Sin embargo, comparte con la caseína β la propiedad de tener zonas predominantemente hidrofílicas e hidrofóbicas bien marcadas y separadas.
Una particularidad de esta caseína es la presencia de una zona con carga neta positiva entre los aminoácidos 20 y 115. Esta zona con carga neta positiva permite la interacción de la caseína con polisacáridos como los carragenanos, que tienen carga negativa. También tiene en la cadena dos grupos de cisteína.
La caseina κ es la única caseína que tiene parte de las moléculas glicosiladas. El grupo glucídico está formado o bien por un trisacárido o bien por un tetrasacárido unido a un resto de treonina, o bien en la treonina que ocupa la posición 131, en la 133, o bien en otra más próxima aún al extremo carboxilo-terminal de la cadena. Dada la presencia de ácido N-acetil neuramínico, este grupo glucídico aporta carga neta a la caseína &kappa.
La caseína κ se rompe fácilmente por proteolisis en el enlace situado entre la fenilalanina 105 y la metionina 106, en una región rica en restos de prolina y probablemente fácilmente accesible.
Cuando esta proteolisis se produce, el fragmento N-terminal 1-105 (para κ-caseína), que es fundamentalmente hidrofóbico, queda unido a las otras caseínas en la micela, mientras que el fragmento C-terminal 106-169 (caseino macropéptido), muy hidrofílico, y en el que está situado el resto glucídico en las moléculas glicosiladas, queda libre en solución.
Estructura de la caseína κ Se resaltan los restos de pprolina. La flecha seńala el punto de corte por la quimosina
La ruptura de la caseína κ produce la desestabilización de la micela, y (a temperaturas por encima de 20ş C) su agregación. Este proceso es el que se produce en la fabricación de la gran mayoría de los quesos.
Caseína γ
Recibe el nombre de caseina γ un conjunto de fragmentos de la caseína β formados por la acción de la plasmina, una proteinasa presente en la leche. En condiciones normales, esta “caseína” representa alrededor del 3% del total de caseínas. Los fragmentos de caseína β más pequeños formados en este proceso proteolítico quedan eln el lactosuero, y reciben el nombre de “fracción proteosa - peptona”.
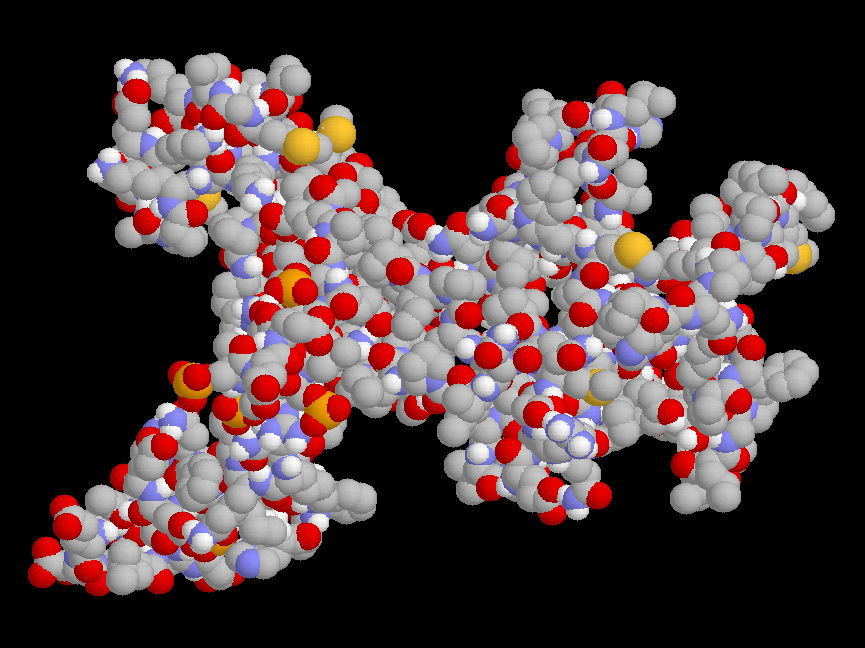
Estructura de las micelas de caseína
Las micelas de caseína son partículas de un tamaño entre 50 y 500 nanometros, formadas por la asociación de moléculas de caseína junto con fosfato cálcico en forma coloidal. El componente “mineral” representa alrededor del 7% del peso de la caseína. La estructura interna de las micelas de caseína, si es que realmente puede hablarse de una estructura organizada, está en discusión. Por una parte, según el modelo más aceptado, las micelas estarían formadas por la agregación de otras partículas menores, las llamadas “submicelas”, unidas entre sí a través de puentes de fosfato y por interacciones hidrofóbicas. Las moléculas individuales de de caseína se unirían dentro de las submicelas fundamentalmente a través de enlaces hidrofóbicos.
Ahora bien, las “submicelas” son probablemente estructuras menos definidas de lo que se ha supuesto hasta el momento, y parece probable que el interior de las micelas tenga una estructura menos organizada, con asociaciones de moléculas de caseína producidas por su interacción con partículas de fosfato cálcico coloidal de tamaño nanométrico. De hecho, el otro modelo de micela de caseína propuesto implica la existencia de nanoclusters de fosfato cálcico distribuido dentro de una partícula más o menos homogénea formada por la asociación de caseínas individuales.
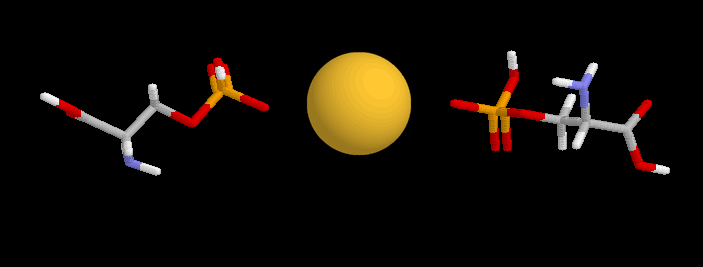
La fosfoserina, que es una pieza esencial en el mantenimiento de la estructura de cualquier modelo de micela de caseína, puede formar puentes con iones de calcio.
Puente de calcio entre dos fosfoserinas
El fosfato de calcio se encuentra en las caseínas como “fosfato coloidal”, en forma no cristalina. Las partículas de fosfato amorfo o coloidal dentro de la caseínas tienen un tamaño del orden de 2,5 nanometros, y están formadas por un núcleo de fosfato cálcico recubierto de una capa de proteína de un espesor de alrededor de 1,5 nanometros, unida al fosfato cálcico mediante puentes a través de las fosfoserinas.
La superficie exterior de la micela sería también cambiante, especialmente sensible a las modificaciones del medio que alteraran la hidratación de las moléculas.
Estabilidad de las micelas de caseína
En condiciones normales de pH y concentración de sales, las micelas de caseínas se encuentra muy hidratadas,
teniendo ligados alrededor de 3,7 gramos de agua por gramo de proteína. La hidrofilicidad de las micelas de caseína se debe a que la
caseína β situá su zona hidrófila preferentemente hacia el exterior, y también lo hace la caseína κ, que se sitúa preferentemente en la superficie de la micela
Las micelas de caseína se desestabilizan fundamentalmente por dos procesos: Por la acidez, y por la proteolisis de la caseína κ.
La acidez tiene dos efectos: En primer lugar, según va bajando el pH se van rompiendo los enlaces entre los grupos fosfato y el ion calcio, al reducirse la ionización de los fosfatos. En segundo lugar, las repulsiones entre las micelas se reducen, al acercarse el pH al punto isoeléctrico de las caseínas. A un pH de alrededor de 4,5 (y a una temperatura superior a 20şC) las caseínas se agregan, formando una cuajada poco mineralizada.
En el tratamiento con quimosina, la caseína κ pierde por proteolisis sus región hidrófila, dirigida hacia el exterior de las micelas. La reducción de la hidrofilicidad facilita la agregación.
A temperaturas bajas, de refrigeración, las fuerzas hidrofóbicas que mantienen unidas a las moléculas de caseína se debilitan, e incluso una parte de la caseína sale de la micela. La gran mayoría permanece, pero unida menos fuertemente. En particular, las fuerzas que actúan sobre la región hidrofóbica de la caseína β se debilitan, haciendo que esta exponga más hacia el exterior su región hidrófila. Esto aumenta la hidratación y voluminosidad de la micela. Como consecuencia, a temperaturas de refrigeración no se produce la agregación de la caseína ni por la acción de la acidez ni por la de la quimosina.
 Volver a Bioquímica de los Alimentos
Volver a Bioquímica de los Alimentos