
Cristales de toxina de Bacillus thuringiensis serovar morrisoni strain T08025. Fotografía de Jim Buckman
|
BIOQUIMICA DE LOS ALIMENTOS Miguel Calvo |
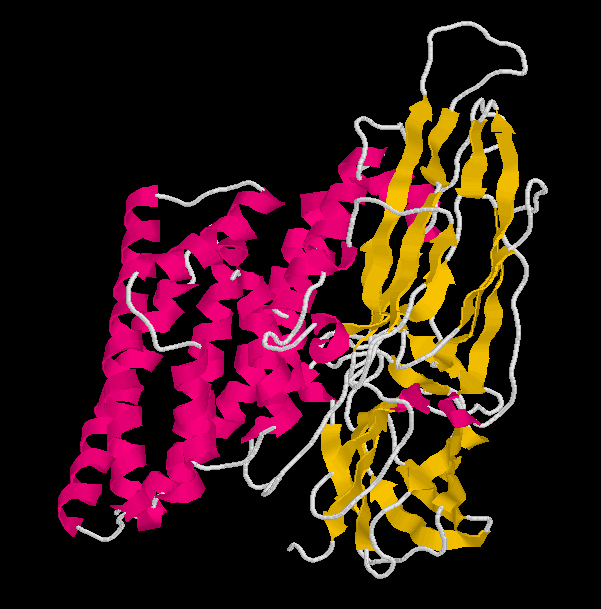

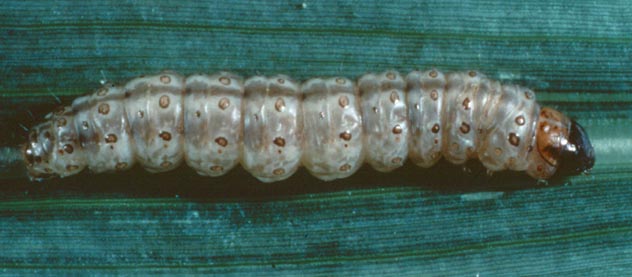

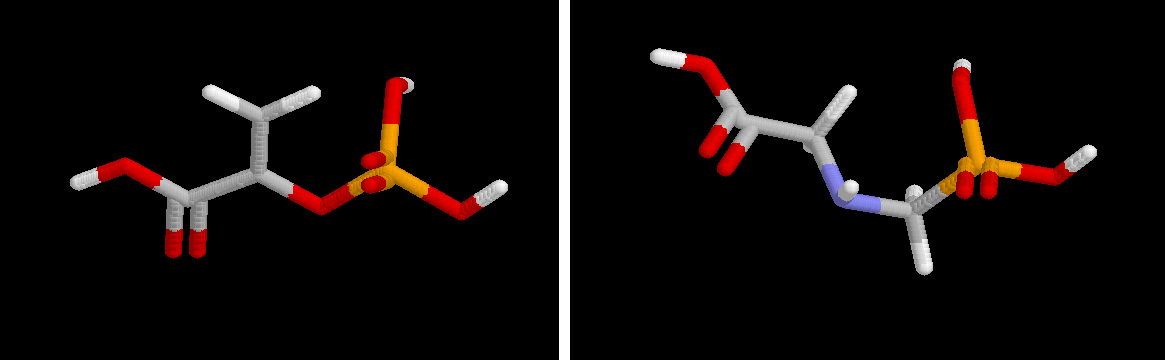




 Volver a Bioquímica de los Alimentos
Volver a: Fundamentos de los alimentos transgénicos
Volver a Bioquímica de los Alimentos
Volver a: Fundamentos de los alimentos transgénicos